Хромосомы человека. Строение хромосом схема строения хромосомы в поздней профазе — метафазе митоза
Хромосомы - это основные структурные элементы клеточного ядра, являющиеся носителями генов, в которых закодирована наследственная информация. Обладая способностью к самовоспроизведению, хромосомы обеспечивают генетическую связь поколений.
Морфология хромосом связана со степенью их спирализации. Например, если в стадии интерфазы (см. Митоз, Мейоз) хромосомы максимально развернуты, т. е. деспирализованы, то с началом деления хромосомы интенсивно спирализуются и укорачиваются. Максимальной спирализации и укорочения хромосомы достигают в стадии метафазы, когда происходит формирование относительно коротких, плотных, интенсивно окрашивающихся основными красителями структур. Эта стадия наиболее удобна для изучения морфологических характеристик хромосом.
Метафазная хромосома состоит из двух продольных субъединиц - хроматид [ выявляет в строении хромосом элементарные нити (так называемые хромонемы, или хромофибриллы) толщиной 200 Å, каждая из которых состоит из двух субъединиц].
Размеры хромосом растений и животных значительно колеблются: от долей микрона до десятков микрон. Средние длины метафазных хромосом человека лежат в пределах 1,5-10 микрон.
Химической основой строения хромосом являются нуклеопротеиды - комплексы (см.) с основными белками - гистонами и протаминами.
Рис. 1. Строение нормальной хромосомы.
А - внешний вид; Б - внутреннее строение: 1-первичная перетяжка; 2 - вторичная перетяжка; 3 -спутник; 4 - центромера.
Индивидуальные хромосомы (рис. 1) различают по локализации первичной перетяжки, т. е. места расположения центромеры (во время митоза и мейоза к этому месту прикрепляются нити веретена, подтягивая ее при этом к полюсу). При утрате центромеры фрагменты хромосом утрачивают способность расходиться при делении. Первичная перетяжка делит хромосомы на 2 плеча. В зависимости от расположения первичной перетяжки хромосомы подразделяют на метацентрические (оба плеча равной или почти равной длины), субметацентрические (плечи неравной длины) и акроцентрические (центромера смещена на конец хромосомы). Помимо первичной, в хромосомах могут встречаться менее выраженные вторичные перетяжки. Небольшой концевой участок хромосом, отделенный вторичной перетяжкой, называют спутником.
Каждый вид организмов характеризуется своим специфическим (по числу, размерам и форме хромосом) так называемым хромосомным набором. Совокупность двойного, или диплоидного, набора хромосом обозначают как кариотип.

Рис. 2. Нормальный хромосомный набор женщины (в правом нижнем углу две X-хромосомы).

Рис. 3. Нормальный хромосомный набор мужчины (в правом нижнем углу - последовательно Х- и Y-хромосомы).
В зрелых , яйцеклетках и содержится одиночный, или гаплоидный, набор хромосом (n), составляющий половину диплоидного набора (2n), присущего хромосомам всех остальных клеток организма. В диплоидном наборе каждая хромосома представлена парой гомологов, один из которых материнского, а другой отцовского происхождения. В большинстве случаев хромосомы каждой пары идентичны по размерам, форме и генному составу. Исключение составляют половые хромосомы, наличие которых определяет развитие организма в мужском или женском направлении. Нормальный хромосомный набор человека состоит из 22 пар аутосом и одной пары половых хромосом. У человека и других млекопитающих женский определяется наличием двух Х-хромосом, а мужской - одной X-и одной Y-хромосомы (рис. 2 и 3). В женских клетках одна из Х-хромосом генетически неактивна и обнаруживается в интерфазном ядре в виде (см.). Изучение хромосом человека в норме и патологии составляет предмет медицинской цитогенетики. Установлено, что отклонения в числе или структуре хромосом от нормы, возникающие в половых! клетках или на ранних этапах дробления оплодотворенной яйцеклетки, вызывают нарушения нормального развития организма, обусловливая в некоторых случаях возникновение части спонтанных абортов, мертворождений, врожденных уродств и аномалий развития после рождения (хромосомные болезни). Примерами хромосомных болезней могут служить болезнь Дауна (лишняя G-хромосома), синдром Клайнфелтера (лишняя Х-хромосома у мужчин) и (отсутствие в кариотипе Y- или одной из Х-хромосом). В медицинской практике хромосомный анализ проводят или прямым методом (на клетках костного мозга), или после кратковременного культивирования клеток вне организма (периферическая кровь, кожа, эмбриональные ткани).
Хромосомы (от греч. chroma - окраска и soma - тело) - нитевидные, самовоспроизводящиеся структурные элементы клеточного ядра, содержащие в линейном порядке факторы наследственности - гены. Хромосомы отчетливо видны в ядре во время деления соматических клеток (митоза) и во время деления (созревания) половых клеток - мейоза (рис. 1). В том и в другом случае хромосомы интенсивно окрашиваются основными красителями, а также видны на неокрашенных цитологических препаратах в фазовом контрасте. В интерфазном ядре хромосомы деспирализованы и не видны в световой микроскоп, так как их поперечные размеры выходят за пределы разрешающей способности светового микроскопа. В это время отдельные участки хромосом в виде тонких нитей диаметром 100-500 Å можно различить при помощи электронного микроскопа. Отдельные не деспирализовавшиеся участки хромосом в интерфазном ядре видны через световой микроскоп как интенсивно красящиеся (гетеропикнотические) участки (хромоцентры).
Хромосомы непрерывно существуют в клеточном ядре, претерпевая цикл обратимой спирализации: митоз-интерфаза-митоз. Основные закономерности строения и поведения хромосом в митозе, мейозе и при оплодотворении одинаковы у всех организмов.
Хромосомная теория наследственности . Впервые хромосомы описали И. Д. Чистяков в 1874 г. и Страсбургер (Е. Strasburger) в 1879 г. В 1901 г. Уилсон (Е. В. Wilson), а в 1902 г. Саттон (W. S. Sutton) обратили внимание на параллелизм в поведении хромосом и менделевских факторов наследственности - генов - в мейозе и при оплодотворении и пришли к выводу, что гены находятся в хромосомах. В 1915-1920 гг. Морган (Т. Н. Morgan) и его сотрудники доказали это положение, локализовали в хромосомах дрозофилы несколько сот генов и создали генетические карты хромосом. Данные о хромосомах, полученные в первой четверти 20 века, легли в основу хромосомной теории наследственности, согласно которой преемственность признаков клеток и организмов в ряду их поколений обеспечивается преемственностью их хромосомах.
Химический состав и ауторепродукция хромосом . В результате цитохимических и биохимических исследований хромосом в 30 и 50-х годах 20 века установлено, что они состоят из постоянных компонентов [ДНК (см. Нуклеиновые кислоты), основных белков (гистонов или протаминов), негистонных белков] и переменных компонентов (РНК и связанного с ней кислого белка). Основу хромосом составляют дезоксирибонуклеопротеидные нити диаметром около 200 Å (рис. 2), которые могут соединяться в пучки диаметром 500 А.
Открытие Уотсоном и Криком (J. D. Watson, F. Н. Crick) в 1953 г. строения молекулы ДНК, механизма ее авторепродукции (редупликации) и нуклеинового кода ДНК и развитие возникшей после этого молекулярной генетики привело к представлению о генах как участках молекулы ДНК. (см. Генетика). Вскрыты закономерности авторепродукции хромосом [Тейлор (J. Н. Taylor) и др., 1957], оказавшиеся аналогичными закономерностям авторепродукции молекул ДНК (полуконсервативная редупликация).
Хромосомный набор - совокупность всех хромосом в клетке. Каждый биологический вид обладает характерным и постоянным набором хромосом, закрепленным в эволюции данного вида. Различают два основных типа наборов хромосом: одиночный, или гаплоидный (в половых клетках животных), обозначаемый n, и двойной, или диплоидный (в соматических клетках, содержащий пары сходных, гомологичных хромосом от матери и отца), обозначаемый 2n.
Наборы хромосом отдельных биологических видов значительно различаются по числу хромосом: от 2 (лошадиная аскарида) до сотен и тысяч (некоторые споровые растения и простейшие). Диплоидные числа хромосом некоторых организмов таковы: человека - 46, гориллы - 48, кошки - 60, крысы - 42, дрозофилы - 8.
Размеры хромосом у разных видов также различны. Длина хромосом (в метафазе митоза) варьирует от 0,2 мк у одних видов до 50 мк у других, а диаметр от 0,2 до 3 мк.
Морфология хромосом хорошо выражена в метафазе митоза. Именно метафазные хромосомы используют для идентификации хромосом. В таких хромосомах хорошо видны обе хроматиды, на которые продольно расщеплена каждая хромосома и центромер (кинетохор, первичная перетяжка), соединяющий хроматиды (рис. 3). Центромер виден как суженный участок, не содержащий хроматина (см.); к нему крепятся нити ахроматинового веретена, благодаря чему центромер определяет движение хромосом к полюсам в митозе и мейозе (рис. 4).
Потеря центромера, например при разрыве хромосомы ионизирующими излучениями или другими мутагенами, приводит к потере способности куска хромосомы, лишенного центромера (ацентрический фрагмент), участвовать в митозе и мейозе и к потере его из ядра. Это может привести к тяжелому повреждению клетки.
Центромер делит тело хромосомы на два плеча. Расположение центромера строго постоянно для каждой хромосомы и определяет три типа хромосом: 1) акроцентрические, или палочкообразные, хромосомы с одним длинным и вторым очень коротким плечом, напоминающим головку; 2) субметацентрические хромосомы с длинными плечами неравной длины; 3) метацентрические хромосомы с плечами одинаковой или почти одинаковой длины (рис. 3, 4, 5 и 7).

Рис. 4. Схема строения хромосом в метафазе митоза после продольного расщепления центромера: А и А1 - сестринские хроматиды; 1 - длинное плечо; 2 - короткое плечо; 3 - вторичная перетяжка; 4- центромер; 5 - волокна веретена.
Характерными чертами морфологии определенных хромосом являются вторичные перетяжки (не обладающие функцией центромера), а также спутники - маленькие участки хромосом, соединенные с остальным ее телом тонкой нитью (рис. 5). Спутничные нити обладают способностью формировать ядрышки. Характерная структура в хромосоме (хромомеры) - утолщения или более плотно спирализованные участки хромосомной нити (хромонемы). Рисунок хромомер специфичен для каждой пары хромосом.

Рис. 5. Схема морфологии хромосомы в анафазе митоза (хроматида. отходящая к полюсу). А - внешний вид хромосомы; Б - внутреннее строение той же хромосомы с двумя составляющими ее хромонемами (полухроматидами): 1 - первичная перетяжка с хромомерами, составляющими центромер; 2 - вторичная перетяжка; 3 - спутник; 4 - нить спутника.
Число хромосом, их размеры и форма на стадии метафазы характерны для каждого вида организмов. Совокупность этих признаков набора хромосом называется кариотипом. Кариотип можно представить в виде схемы, называемой идиограммой (см. ниже хромосомы человека).
Половые хромосомы . Гены, детерминирующие пол, локализованы в специальной паре хромосом - половых хромосомах (млекопитающие, человек); в других случаях иол определяется соотношением числа половых хромосом и всех остальных, называемых аутосомами (дрозофила). У человека, как и у других млекопитающих, женский пол определяется двумя одинаковыми хромосомами, обозначаемыми как Х-хромосомы, мужской пол определяется парой гетероморфных хромосом: Х и Y. В результате редукционного деления (мейоза) при созревании ооцитов (см. Овогенез) у женщин все яйца содержат по одной Х-хромосоме. У мужчин в результате редукционного деления (созревания) сперматоцитов половина спермиев содержит Х-хромосому, а другая половина Y-хромосому. Пол ребенка определяется случайным оплодотворением яйцеклетки спермием, несущим Х- или Y-хромосому. В результате возникает зародыш женского (XX) или мужского (XY) пола. В интерфазном ядре у женщин одна из Х-хромосом видна как глыбка компактного полового хроматина.
Функционирование хромосом и метаболизм ядра . Хромосомная ДНК является матрицей для синтеза специфических молекул информационной РНК. Этот синтез происходит тогда, когда данный участок хромосомы деспирализован. Примерами локальной активации хромосом служат: образование деспирализованных петель хромосом в ооцитах птиц, амфибий, рыб (так называемые Х-ламповые щетки) и вздутий (пуффов) определенных локусов хромосом в многонитчатых (политенных) хромосомах слюнных желез и других секреторных органов двукрылых насекомых (рис. 6). Примером инактивации целой хромосомы, т. е. выключения ее из метаболизма данной клетки, является образование одной из Х-хромосом компактного тела полового хроматина.
Рис. 6. Политенные хромосомы двукрылого насекомого Acriscotopus lucidus: А и Б - участок, ограниченный пунктирными линиями, в состоянии интенсивного функционирования (пуфф); В - тот же участок в нефункционирующем состоянии. Цифрами обозначены отдельные локусы хромосом (хромомеры).
Рис. 7. Хромосомный набор в культуре лейкоцитов периферической крови мужчины (2n=46).
Вскрытие механизмов функционирования политенных хромосом типа ламповых щеток и других типов спирализации и деспирализации хромосом имеет решающее значение для понимания обратимой дифференциальной активации генов.
Хромосомы человека . В 1922 г. Пейнтер (Т. S. Painter) установил диплоидное число хромосом человека (в сперматогониях), равное 48. В 1956 г. Тио и Леван (Н. J. Tjio, A. Levan) использовали комплекс новых методов исследования хромосом человека: культуру клеток; исследование хромосом без гистологических срезов на тотальных препаратах клеток; колхицин, приводящий к остановке митозов на стадии метафазы и накоплению таких метафаз; фитогемагглютинин, стимулирующий вступление клеток в митоз; обработку метафазных клеток гипотоническим солевым раствором. Все это позволило уточнить диплоидное число хромосом у человека (оно оказалось равным 46) и дать описание кариотипа человека. В 1960 г. в Денвере (США) международная комиссия разработала номенклатуру хромосом человека. Согласно предложениям комиссии, термин «кариотип» следует применять к систематизированному набору хромосом единичной клетки (рис. 7 и 8). Термин «идиотрамма» сохраняется для представления о наборе хромосом в виде диаграммы, построенной на основании измерений и описания морфологии хромосом нескольких клеток.
Хромосомы человека пронумерованы (отчасти серийно) от 1 до 22 в соответствии с особенностями морфологии, допускающими их идентификацию. Половые хромосомы не имеют номеров и обозначаются как Х и Y (рис. 8).
Обнаружена связь ряда заболеваний и врожденных дефектов в развитии человека с изменениями в числе и структуре его хромосом. (см. Наследственность).
См. также Цитогенетические исследования.
Все эти достижения создали прочную базу для развития цитогенетики человека.
Рис. 1. Хромосомы: А - на стадии анафазы митоза в микроспороцитах трилистника; Б - на стадии метафазы первого деления мейоза в материнских клетках пыльцы у традесканции. В обоих случаях видно спиральное строение хромосом.
Рис. 2. Элементарные хромосомные нити с диаметром 100 Å (ДНК + гистон) из интерфазных ядер вилочковой железы теленка (электронная микроскопия): А - изолированные из ядер нити; Б - тонкий срез через пленку того же препарата.
Рис. 3. Хромосомный набор Vicia faba (конские бобы) в стадии метафазы.
Рис. 8. Хромосомы того же, что на рис. 7, набора, систематизированные согласно денверовской номенклатуре в пары гомологов (кариотип).
Энциклопедичный YouTube
1 / 5
Хромосомы, хроматиды, хроматин и т.п.
Гены, ДНК и хромосомы
Тайны X-хромосомы
Биология в картинках: Строение хромосомы (Вып. 70)
Является ли Y-хромосома исчезающей | Озвучка DeeAFilm
Субтитры
Перед погружением в механизм деления клеток, я думаю, будет полезно поговорить о лексике, связанной с ДНК. Есть много слов, и некоторые из них сходны по звучанию друг с другом. Они могут сбивать с толку. Для начала я бы хотел поговорить о том, как ДНК генерирует больше ДНК, создаёт свои копии, или о том, как она вообще делает белки. Мы уже говорили об этом в ролике о ДНК. Давайте я нарисую небольшой участок ДНК. У меня есть A, Г, T, пусть у меня Есть два Т и потом два Ц. Такой небольшой участок. Он продолжается вот так. Конечно, это двойная спираль. Каждой букве соответствует своя. Я нарисую их этим цветом. Итак, A соответствует T, Г соответствует Ц, (точнее Г образует водородные связи с Ц), T - с A, T - с A, Ц - с Г, Ц - с Г. Вся эта спираль тянется, допустим, в этом направлении. Итак, есть пара различных процессов, которые эта ДНК должна осуществить. Один из них связан с клетками вашего тела - необходимо произвести больше клеток вашей кожи. Ваша ДНК должна скопировать себя. Этот процесс называется репликацией. Вы реплицируете ДНК. Я покажу вам репликацию. Как эта ДНК может скопировать себя? Это одна из самых замечательных особенностей структуры ДНК. Репликация. Я делаю общее упрощение, но идея заключается в том, что две цепи ДНК разделяются, и это происходит не само по себе. Этому способствует масса белков и ферментов, но в деталях я буду рассказывать о микробиологии в другом ролике. Итак, эти цепи отделяются друг от друга. Я перенесу цепь сюда. Они отделяются друг от друга. Я возьму другую цепь. Эта слишком большая. Эта цепь будет выглядеть как-то так. Они отделяются друг от друга. Что же может произойти после этого? Я удалю лишние фрагменты здесь и здесь. Итак, вот наша двойная спираль. Они все были связаны. Это пары оснований. Теперь они отделяются друг от друга. Что может делать каждая из них после разделения? Они теперь могут стать матрицей друг для друга. Смотрите… Если эта цепь находится сама по себе, сейчас, неожиданно может прийти тиминовое основание и присоединится здесь, и эти нуклеотиды начнут выстраиваться в линию. Тимин и цитозин, и потом аденин, аденин, гуанин, гуанин. И так продолжаться. И тогда, в этой другой части, на зелёной цепи, которая была до этого прикреплена к этой голубой, будет происходить то же самое. Будет аденин, гуанин, тимин, тимин, цитозин, цитозин. Что произошло только что? Разделением и привлечением комплементарных оснований, мы создали копию этой молекулы. Мы займёмся микробиологией этого в будущем, это только для общего представления о том, как ДНК копирует себя. Особенно, когда мы рассматриваем митоз и мейоз, я могу сказать: «Это стадия, где происходит репликация». Теперь, другой процесс, о котором вы ещё много услышите. Я говорил о нём в ролике о ДНК. Это транскрипция. В ролике о ДНК я не уделял много внимания тому, как ДНК удваивает сама себя, но одна из великолепных особенностей устройства двойной цепи - это лёгкая возможность самоудвоения. Вы просто разделяете 2 полоски, 2 спирали, а потом они становятся матрицей для другой цепи, и тогда появляется копия. Теперь транскрипция. Это то, что должно произойти с ДНК для того, чтобы образовались белки, но транскрипция - это промежуточная стадия. Это стадия, когда вы переходите от ДНК к мРНК. Тогда эта мРНК покидает ядро клетки и направляется к рибосомам. Я буду говорить об этом через несколько секунд. Итак, мы можем сделать то же самое. Эти цепи опять в ходе транскрипции разделяются. Одна отделяется сюда, а другая отделяется... а другая будет отделятся вот сюда. Прекрасно. Может быть имеет смысл использовать только одну половину цепи - я удалю одну. Вот таким образом. Мы собираемся транскрибировать зелёную часть. Вот она. Всё это я удалю. Не тот цвет. Итак, я удаляю всё это. Что произойдёт, если вместо нуклеотидов дезоксирибонуклеиновой кислоты, которые образуют пары с этой цепью ДНК, у вас есть рибонуклеиновая кислота, или РНК, образующая пары. Изображу РНК пурпурным цветом. РНК будет образовывать пары с ДНК. Тимин, находящийся в ДНК, будет образовывать пару с аденином. Гуанин, теперь, когда мы говорим о РНК, вместо тимина у нас будет урацил, урацил, цитозин, цитозин. И это будет продолжаться. Это мРНК. Информационная РНК. Теперь она отделяется. Эта мРНК отделяется и покидает ядро. Она покидает ядро, и тогда происходит трансляция. Трансляция. Запишем этот термин. Трансляция. Это идёт от мРНК... В ролике о ДНК у меня была маленькая тРНК. Транспортная РНК была как бы грузовиком, перевозящим аминокислоты к мРНК. Всё это происходит в части клетки, называемой рибосомой. Трансляция происходит от мРНК к белку. Мы видели, как это происходит. Итак, от мРНК к белку. У вас есть эта цепь - я сделаю копию. Скопирую всю цепь сразу. Эта цепь отделяется, покидает ядро, и тогда у вас есть эти маленькие грузовики тРНК, которые, собственно, и, так сказать, подъезжают. Итак, допустим, у меня есть тРНК. Давайте посмотрим, аденин, аденин, гуанин и гуанин. Это РНК. Это кодон. Кодон имеет 3 пары оснований и прикреплённую к нему аминокислоту. У вас есть некоторые другие части тРНК. Скажем, урацил, цитозин, аденин. И прикреплённая к нему другая аминокислота. Тогда аминокислоты соединяются и образуют длинную цепь аминокислот, которая является белком. Белки образуют эти странные сложные формы. Чтобы убедиться, что вы поняли. Мы начнём с ДНК. Если мы производим копии ДНК - это репликация. Вы реплицируете ДНК. Итак, если мы производим копии ДНК - это репликация. Если вы начинаете с ДНК и создаёте мРНК с матрицы ДНК, то это транскрипция. Запишем. "Транскрипция" . То есть вы транскрибируете информацию с одной формы на другую - транскрипция. Теперь, когда мРНК покидает ядро клетки… Я нарисую клетку, чтобы обратить на это внимание. Мы займёмся структурой клетки в будущем. Если это целая клетка, ядро - это центр. Это место, где находятся все ДНК, все репликации и транскрипции происходят здесь. Затем мРНК покидает ядро, и тогда в рибосомах, которые мы более подробно обсудим в будущем, происходит трансляция и формируется белок. Итак, от мРНК к белку - это трансляция. Вы транслируете с генетического кода, в так называемый белковый код. Итак, это и есть трансляция. Это именно те слова, которые обычно используются для описания этих процессов. Убедитесь, что вы правильно их используете, называя различные процессы. Теперь другая часть терминологии ДНК. Когда я впервые встретился с ней, я решил, что она чрезвычайно сбивает с толку. Это слово «хромосома». Запишу слова здесь - вы сами можете оценить, как они сбивают с толку: хромосома, хроматин и хроматида. Хроматида. Итак, хромосома, мы уже говорили о ней. У вас может быть цепь ДНК. Это двойная спираль. Эта цепь, если я увеличу её, - на самом деле две разных цепи. Они имеют соединённые пары оснований. Я только что нарисовал пары оснований, соединённые вместе. Я хочу, чтобы было ясно: я нарисовал эту небольшую зелёную линию здесь. Это двойная спираль. Она оборачивается вокруг белков, которые называются гистонами. Гистоны. Пусть она оборачивается вот так и как-то так, а потом как-нибудь так. Здесь у вас есть вещества, называемые гистонами, которые являются белками. Нарисуем их вот таким образом. Вот так. Это структура, то есть ДНК в комбинации с белками, которые её структурируют, заставляя оборачиваться вокруг дальше и дальше. В конечном счёте, в зависимости от стадии жизни клетки, будут образовываться различные структуры. И когда вы говорите о нуклеиновой кислоте, которая является ДНК, и объединяете её с белками, то вы говорите о хроматине. Значит, хроматин - это ДНК плюс структурные белки, которые придают ДНК форму. Структурные белки. Идея хроматина была впервые использована из-за того, что люди видели, когда смотрели на клетку… Помните? Каждый раз я рисовал клеточное ядро определённым образом. Скажем, так. Это ядро клетки. Я рисовал очень хорошо различимые структуры. Это одна, это другая. Может быть, она короче, и у неё есть гомологичная хромосома. Я нарисовал хромосомы, так? И каждая из этих хромосом, как я уже показывал в прошлом видео, - по существу - длинные структуры ДНК, длинные цепи ДНК, плотно обёрнутые друг вокруг друга. Я рисовал это как-то так. Если мы увеличим, то увидим одну цепь, и она действительно обёрнута вокруг себя подобно этому. Это её гомологичная хромосома. Вспомните, в ролике, посвящённом изменчивости, я говорил о гомологичной хромосоме, которая кодирует те же гены, но другую их версию. Синий - от папы, а красный - от мамы, но они по существу кодируют те же гены. Итак, это одна цепь, которую я получил от папы с ДНК этой структуры, мы называем её хромосомой. Итак, хромосома. Я хочу, чтобы это было ясно, ДНК принимает эту форму только на определённых жизненных стадиях, когда она воспроизводит сама себя, т.е. реплицируется. Точнее не так… Когда клетка делится. Перед тем как клетка становится способной к делению, ДНК принимает эту хорошо определённую форму. Большую часть жизни клетки, когда ДНК делает свою работу, когда она создаёт белки, то есть белки транскрибируются и транслируются с ДНК, она не сворачивается таким образом. Если бы она была свёрнута, для репликационной и транскрипционной системы было бы затруднительно проникнуть к ДНК, произвести белки и делать что-то ещё. Обычно ДНК… Давайте я ещё раз нарисую ядро. Чаще всего вы даже не можете увидеть её в обычный световой микроскоп. Она настолько тонкая, что вся спираль ДНК полностью распределена в ядре. Я рисую это здесь, другая может быть здесь. А потом у вас есть более короткая цепь, типа этой. Вы даже не можете её увидеть. Она не находится в этой, хорошо определённой структуре. Обычно это выглядит таким образом. Пусть будет ещё такая короткая цепь. Вы можете увидеть только подобный беспорядок, состоящий из путаницы комбинаций ДНК и белков. Это то, что люди в общем-то и называют хроматином. Это нужно записать. "Хроматин" Таким образом, слова могут быть очень неоднозначны и очень запутанны, но общее использование, когда вы говорите о хорошо определённой одной цепи ДНК, вот таким образом хорошо определённой структуры, то это хромосома. Понятие "хроматин" может относиться либо к структуре типа хромосомы, комбинации ДНК и белков, структурирующих ее, либо к беспорядку множества хромосом, в которых есть ДНК. То есть из множества хромосом и белков, перемешанных вместе. Я хочу, чтобы это было понятно. Теперь следующее слово. Что такое хроматида? На всякий случай, если я ещё не сделал этого… Я не помню, помечал ли я это. Эти белки, которые обеспечивают структуру хроматина или составляют хроматин, а также обеспечивают структуру называются "гистонами". Есть различные типы, которые обеспечивают структуру на различных уровнях, мы ещё рассмотрим их детально. Итак, что такое хроматида? Когда ДНК реплицируется… Скажем, это была моя ДНК, она находится в нормальном состоянии. Одна версия - от папы, одна версия - от мамы. Теперь она реплицируется. Версия от папы сначала выглядит так. Это большая цепь ДНК. Она создаёт другую версию себя, идентичную, если система работает правильно, и эта идентичная часть выглядит так. Они изначально прикреплены друг к другу. Они прикреплены друг к другу в месте, называемом центромерой. Теперь, несмотря на то что у меня здесь 2 цепи, скрепленные вместе. Две одинаковые цепи. Одна цепь здесь, одна тут… Хотя давайте я изображу иначе. В принципе это можно изобразить множеством разных способов. Это одна цепь здесь, и вот другая цепь тут. То есть у нас имеются 2 копии. Они кодируют абсолютно одинаковую ДНК. Так вот. Они идентичны, поэтому я всё ещё называю это хромосомой. Запишем это тоже. Всё это вместе называется хромосомой, но теперь каждая отдельная копия называется хроматидой. Итак, это одна хроматида и это другая. Иногда их называют сестринскими хроматидами. Также их можно назвать хроматидами-близнецами, потому что у них одна и та же генетическая информация. Итак, эта хромосома имеет 2 хроматиды. Теперь перед репликацией или перед удвоением ДНК вы можете сказать, что эта хромосома вот здесь имеет одну хроматиду. Вы можете называть это хроматидой, но это не обязательно. Люди начинают говорить о хроматидах тогда, когда две из них присутствуют в хромосоме. Мы узнаем, что в митозе и мейозе эти 2 хроматиды разделяются. Когда они разделяются, тут же цепь ДНК, которую вы однажды называли хроматидой, теперь вы будете называть отдельной хромосомой. Итак, это одна из них, и вот другая, которая могла отделиться в этом направлении. Обведу эту зелёным. Итак, эта может отойти в эту сторону, а эта, которую я обвёл оранжевым, например, в эту … Теперь, когда они отделены и больше не связаны центромерой, то, что мы изначально называли одной хромосомой с двумя хроматидами, теперь вы называете двумя отдельными хромосомами. Или можно сказать, что теперь у вас есть две отдельные хромосомы, каждая из которых состоит из одной хроматиды. Я надеюсь, что это немного проясняет значение терминов, связанных с ДНК. Я всегда находил их довольно запутанными, но они будут полезным инструментом, когда мы начнём митоз и мейоз и я буду говорить о том, что хромосома становится хроматидой. Вы будете спрашивать, как одна хромосома стала двумя хромосомами, и как хроматида стала хромосомой. Всё это вращается вокруг лексики. Я бы выбрал другую, вместо того чтобы называть это хромосомой и каждую из этих отдельными хромосомами, но так решили называть за нас. Возможно, вам интересно узнать, откуда это слово - «хромо». Может быть, вы знаете старую плёнку «Кодак», которая называлась «хромо цвет». В принципе «хромо» означает «цвет». Я думаю, оно происходит от греческого слова «цвет». Когда люди первый раз стали рассматривать ядро клетки, они использовали краситель, и то, что мы называем хромосомами, окрашивалось красителем. И мы могли видеть это в световой микроскоп. Часть «сома» происходит от слова «сома», обозначающего «тело», то есть мы получаем окрашенное тело. Так появилось слово «хромосома». Хроматин также окрашивается… Надеюсь, это немного проясняет понятия «хроматида», «хромосома», «хроматин», и теперь мы подготовлены к изучению митоза и мейоза.
История открытия хромосом
Первые описания хромосом появились в статьях и книгах разных авторов в 70-х годах XIX века, и приоритет открытия хромосом отдают разным людям. Среди них такие имена, как И. Д. Чистяков (1873), А. Шнейдер (1873), Э. Страсбургер (1875), О. Бючли (1876) и другие . Чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем - немецкого анатома В. Флеминга , который в своей фундаментальной книге «Zellsubstanz, Kern und Zelltheilung» (нем.) собрал и упорядочил сведения о них, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Г. Вальдейером в 1888 году. «Хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами .
После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы при мейозе и оплодотворении ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902-1903 годах У. Сеттон (Walter Sutton ) независимо друг от друга выдвинули гипотезу о генетической роли хромосом .
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине .
Морфология метафазных хромосом
На стадии метафазы митоза хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации . У метафазных хромосом сестринские хроматиды соединены в районе первичной перетяжки , называемой центромерой . Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора - сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления - движителям хромосомы в митозе . Центромера делит хромосомы на две части, называемые плечами . У большинства видов короткое плечо хромосомы обозначают буквой p , длинное плечо - буквой q . Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин . Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрические хромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует .
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка , которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают различной длины и могут располагаться в различных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы , содержащие многократные повторы генов, кодирующих рибосомные РНК . У человека вторичные перетяжки, содержащие рибосомные гены, находятся в коротких плечах акроцентрических хромосом, они отделяют от основного тела хромосомы небольшие хромосомные сегменты, называемые спутниками . Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) - без ДНК).
Дифференциальная окраска метафазных хромосом
При монохромном окрашивании хромосом (ацето-кармином, ацето-орсеином, окрашиванием по Фёльгену или Романовскому-Гимзе) можно идентифицировать число и размеры хромосом; их форму, определяемую прежде всего положением центромер, наличием вторичных перетяжек, спутников. В подавляющем числе случаев для идентификации индивидуальных хромосом в хромосомном наборе этих признаков недостаточно. Кроме того, монохромно окрашенные хромосомы часто очень похожи у представителей разных видов. Дифференциальное окрашивание хромосом, различные методики которого были разработаны в начале 70-х годов XX века, снабдило цитогенетиков мощнейшим инструментом для идентификации как индивидуальных хромосом в целом, так и их частей, облегчив тем самым процедуру анализа генома .
Методы дифференциального окрашивания делятся на две основные группы:
Уровни компактизации хромосомной ДНК
Основу хромосомы составляет линейная макромолекула ДНК значительной длины. В молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований . Суммарная длина ДНК из одной клетки человека составляет величину порядка двух метров. При этом типичное ядро клетки человека, которое можно увидеть только при помощи микроскопа, занимает объём около 110 мкм³, а митотическая хромосома человека в среднем не превышает 5 - 6 мкм. Подобная компактизация генетического материала возможна благодаря наличию у эукариот высокоорганизованной системы укладки молекул ДНК как в интерфазном ядре, так и в митотической хромосоме. Надо отметить, что у эукариот в пролиферирующих клетках осуществляется постоянное закономерное изменение степени компактизации хромосом. Перед митозом хромосомная ДНК компактизуется в 10 5 раз по сравнению с линейной длиной ДНК, что необходимо для успешной сегрегации хромосом в дочерние клетки, в то время как в интерфазном ядре для успешного протекания процессов транскрипции и репликации хромосоме необходимо декомпактизоваться . При этом ДНК в ядре никогда не бывает полностью вытянутой и всегда в той или иной степени упакована. Так, расчётное уменьшение размера между хромосомой в интерфазе и хромосомой в митозе составляет всего примерно 2 раза у дрожжей и 4 - 50 раз у человека .
Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи считают уровень так называемой хромонемы , толщина которой составляет около 0,1 - 0,3 мкм . В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Хромосомные аномалии
Анеуплоидия
При анеуплоидии происходит изменение числа хромосом в кариотипе, при котором общее число хромосом не кратно гаплоидному хромосомному набору n . В случае утраты одной хромосомы из пары гомологичных хромосом мутантов называют моносомиками , в случае одной дополнительной хромосомы мутантов с тремя гомологичными хромосомами называют трисомиками , в случае утраты одной пары гомологов - нуллисомиками . Анеуплоидия по аутосомным хромосомам всегда вызывает значительные нарушения развития, являясь основной причиной спонтанных абортов у человека . Одной из самых известных анеуплоидий у человека является трисомия по хромосоме 21, которая приводит к развитию синдрома Дауна . Анеуплоидия характерна для опухолевых клеток, особенно для клеток сóлидных опухолей .
Полиплоидия
Изменение числа хромосом, кратное гаплоидному набору хромосом (n ), называется полиплоидией. Полиплоидия широко и неравномерно распространена в природе. Известны полиплоидные эукариотические микроорганизмы - грибы и водоросли , часто встречаются полиплоиды среди цветковых, но не среди голосемянных растений. Полиплоидия клеток всего организма у многоклеточных животных редка, хотя у них часто встречается эндополиплоидия некоторых дифференцированных тканей, например, печени у млекопитающих, а также тканей кишечника, слюнных желёз, мальпигиевых сосудов ряда насекомых .
Хромосомные перестройки
Хромосомные перестройки (хромосомные аберрации) - это мутации, нарушающие структуру хромосом. Они могут возникнуть в соматических и зародышевых клетках спонтанно или в результате внешних воздействий (ионизирующее излучение, химические мутагены, вирусная инфекция и др.). В результате хромосомной перестройки может быть утрачен или, наоборот, удвоен фрагмент хромосомы (делеция и дупликация , соответственно); участок хромосомы может быть перенесён на другую хромосому (транслокация) или он может изменить свою ориентацию в составе хромосомы на 180° (инверсия). Существуют и другие хромосомные перестройки.
Необычные типы хромосом
Микрохромосомы
B-хромосомы
B-хромосомы - это добавочные хромосомы, которые имеются в кариотипе только у отдельных особей в популяции. Они часто встречаются у растений , описаны у грибов , насекомых и животных . Некоторые В-хромосомы содержат гены, часто это гены рРНК , однако не ясно, насколько эти гены функциональны. Наличие В-хромосом может влиять на биологические характеристики организмов, особенно у растений, где их наличие ассоциируется с пониженной жизнеспособностью. Предполагается, что В-хромосомы постепенно утрачиваются в соматических клетках в результате нерегулярности их наследования .
Голоцентрические хромосомы
Голоцентрические хромосомы не имеют первичной перетяжки, они имеют так называемый диффузный кинетохор, поэтому во время митоза микротрубочки веретена деления прикрепляются по всей длине хромосомы. Во время расхождения хроматид к полюсам деления у голоцентрических хромосом они идут к полюсам параллельно друг другу, в то время как у моноцентрической хромосомы кинетохор опережает остальные части хромосомы, что приводит к характерной V-образной форме расходящихся хроматид на стадии анафазы. При фрагментации хромосом, например, в результате воздействия ионизирующего излучения, фрагменты голоцентрических хромосом расходятся к полюсам упорядоченно, а не содержащие центромеры фрагменты моноцентрических хромосом распределяются между дочерними клетками случайным образом и могут быть утрачены .
Голоцентрические хромосомы встречаются у протист , растений и животных. Голоцентрическими хромосомами обладает нематода C. elegans .
Гигантские формы хромосом
Политенные хромосомы
Политенные хромосомы - это гигантские скопления объединённых хроматид, возникающие в некоторых типах специализированных клеток. Впервые описаны Е.Бальбиани (Edouard-Gerard Balbiani ) в году в клетках слюнных желёз мотыля (Chironomus ), однако их цитогенетическая роль была выявлена позднее в 30-х годах XX века Костовым, Т. Пэйнтером, Э. Хайцем и Г. Бауером (Hans Bauer ). Политенные хромосомы обнаружены также в клетках слюнных желёз, кишечника , трахей , жирового тела и мальпигиевых сосудов личинок двукрылых .
Хромосомы типа ламповых щёток
Хромосомы типа ламповых щёток - это гигантская форма хромосом, которая возникает в мейотических женских клетках на стадии диплотены профазы I у некоторых животных, в частности, у некоторых земноводных и птиц . Эти хромосомы являются крайне транскрипционно активными и наблюдаются в растущих ооцитах тогда, когда процессы синтеза РНК , приводящие к образованию желтка , наиболее интенсивны. В настоящее время известно 45 видов животных, в развивающихся ооцитах которых можно наблюдать такие хромосомы. Хромосомы типа ламповых щёток не образуются в ооцитах млекопитающих .
Впервые хромосомы типа ламповых щёток были описаны В. Флеммингом в 1882 году . Название «хромосомы типа ламповых щёток» было предложено немецким эмбриологом И. Рюккертом (J.Rϋckert ) в 1892 году .
По длине хромосомы типа ламповых щёток превышают политенные хромосомы. Например, общая длина хромосомного набора в ооцитах некоторых хвостатых амфибий достигает 5900 мкм.
Бактериальные хромосомы
Имеются данные о наличии у бактерий белков, связанных с ДНК нуклеоида , но гистонов у них не обнаружено.
Хромосомы человека
В каждой ядросодержащей соматической клетке человека содержится 23 пары линейных хромосом, а также многочисленные копии митохондриальной ДНК . В нижеприведённой таблице показано число генов и оснований в хромосомах человека.
| Хромосома | Всего оснований | Количество генов | Количество белок-кодирующих генов |
|---|---|---|---|
| 249250621 | 3511 | 2076 | |
| 243199373 | 2368 | 1329 | |
| 198022430 | 1926 | 1077 | |
| 191154276 | 1444 | 767 | |
| 180915260 | 1633 | 896 | |
| 171115067 | 2057 | 1051 | |
| 159138663 | 1882 | 979 | |
| 146364022 | 1315 | 702 | |
| 141213431 | 1534 | 823 | |
| 135534747 | 1391 | 774 | |
| 135006516 | 2168 | 1914 | |
| 133851895 | 1714 | 1068 | |
| 115169878 | 720 | 331 | |
| 107349540 | 1532 | 862 | |
| 102531392 | 1249 | 615 | |
| 90354753 | 1326 | 883 | |
| 81195210 | 1773 | 1209 | |
| 78077248 | 557 | 289 | |
| 59128983 | 2066 | 1492 | |
| 63025520 | 891 | 561 | |
| 48129895 | 450 | 246 | |
| 51304566 | 855 | 507 | |
| X-хромосома | 155270560 | 1672 | 837 |
| Y-хромосома | 59373566 | 429 | 76 |
| Всего | 3 079 843 747 | 36463 |
См. также
Примечания
- Тарантул В. З. Толковый биотехнологический словарь. - М. : Языки славянских культур, 2009. - 936 с. - 400 экз. - ISBN 978-5-9551-0342-6 .
- Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. - М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. - Т. I. - С. 309-336. - 808 с. -
Хромосомы - самовоспроизводящиеся структуры клеточного ядра. Как у прокариотических, так и у эукариотических организмов гены располагаются группами на отдельных молекулах ДНК, которые при участии белков и других макромолекул клеток организуются в хромосомы. Зрелые клетки зародышевой линии (гаметы - яйцеклетки, спермии) многоклеточных организмов содержат по одному (гаплоидному) набору хромосом организма.
После того как к полюсам отойдут полные наборы хроматид, их называют хромосомами . Хромосомы - это структуры в ядре клеток эукариот, которые пространственно и функционально организовывают ДНК в геноме индивидуумов.
Химический состав хромосом. Хромосома представляет собой дезоксирибонуклеопротеид (ДНП), то есть комплекс, образованный из одной непрерывной двухцепочечной молекулы ДНК и белков (гистонов и негистонов). В состав хромосом входят также липиды и минеральные вещества (например, ионы Ca 2+ , Mg 2+).
Каждая хромосома – сложное надмолекулярное образование , сформированное в результате компактизации хроматина.
Строение хромосом. В большинстве случаев хромосомы хорошо видны лишь в делящихся клетках начиная со стадии метафазы, когда их можно видеть даже в световой микроскоп. В этот период удается определить количество хромосом в ядре, их размеры, форму и строение. Именно такие хромосомы называют метафазными. Интерфазные хромосомы часто называют просто хроматином .
Число хромосом обычно постоянно для всех клеток особи любого вида растений, животных и человека. Но у разных видов количество хромосом неодинаково (от двух до нескольких сотен). Наименьшее число хромосом имеет лошадиная аскарида, наибольшее встречается у простейших и папоротников, для которых характерны высокие уровни полиплоидии. Обычно диплоидные наборы содержат от одного до нескольких десятков хромосом.
Количество хромосом в ядре не связано с уровнем эволюционного развития живых организмов. У многих примитивных форм оно велико, например, в ядрах некоторых видов простейших содержатся сотни хромосом, тогда как у шимпанзе их всего только 48.
Каждая хромосома, образованная одной молекулой ДНК, представляет собой удлиненную палочковидную структуру – хроматиду , имеющую два «плеча», разделенных первичной перетяжкой, или центромерой. Метафазная хромосома состоит из двух соединенных центромерой сестринских хроматид, каждая из которых содержит одну молекулу ДНК, уложенную в виде спирали.
Центромера – это небольшое фибриллярное тельце, осуществляющее первичную перетяжку хромосомы. Она является важнейшей частью хромосомы, так как определяет ее движение. Центромеру, к которой прикрепляются нити веретена во время деления (при митозе и мейозе), называют кинетохором (от греч. kinetos – подвижный и choros – место). Он контролирует движение расходящихся хромосом при делении клетки. Хромосома, лишенная центромеры, не способна совершать упорядоченное движение и может потеряться.
Обычно центромера хромосомы занимает определенное место, и это является одним из видовых признаков, по которому различают хромосомы. Изменение положения центромеры в той или иной хромосоме служит показателем хромосомных перестроек. Плечи хромосом оканчиваются участками, не способными соединяться с другими хромосомами или их фрагментами. Эти концевые участки хромосом называют теломерами . Теломеры предохраняют концы хромосом от слипания и тем самым обеспечивают сохранение их целостности. За открытие механизма защиты хромосом теломерами и ферментом теломеразой американские ученые Э. Блекберн, К. Грейдер и Д. Шостак в 2009 году были удостоены Нобелевской премии в области медицины и физиологии. Концы хромосом нередко обогащены гетерохроматином.

В зависимости от расположения центромеры определяют три основных вида хромосом: равноплечие (плечи равной длины), неравноплечие (с плечами разной длины) и палочковидные (с одним, очень длинным и другим, очень коротким, едва заметным плечом). Некоторые хромосомы имеют не только одну центромеру, но еще и вторичную перетяжку, не связанную с прикреплением нити веретена при делении. Этот участок – ядрышковый организатор , выполняющий функцию синтеза ядрышка в ядре.
Репликация хромосом
Важным свойством хромосом является их способность к удвоению (самовоспроизведению). Обычно удвоение хромосом предшествует делению клетки. В основе удвоения хромосом лежит процесс репликации (от лат. replicatio – повторение) макромолекул ДНК, обеспечивающий точное копирование генетической информации и передачу ее от поколения к поколению. Удвоение хромосом – это сложный процесс, включающий в себя не только репликацию гигантских молекул ДНК, но также синтез связанных с ДНК хромосомных белков. Конечным этапом является упаковка ДНК и белков в особые комплексы, образующие хромосому. В результате репликации вместо одной материнской хромосомы появляются две идентичные ей дочерние хромосомы.
Функция хромосом заключается:
- в хранении наследственной информации. Хромосомы являются носителями генетической информации;
- передаче наследственной информации. Наследственная информация передается путем репликации молекулы ДНК;
- реализации наследственной информации. Благодаря воспроизводству того или иного типа и-РНК и, соответственно, того или иного типа белка осуществляется контроль над всеми процессами жизнедеятельности клетки и всего организма.
Таким образом, хромосомы с заключенными в них генами обусловливают непрерывный ряд воспроизведения.
Хромосомы осуществляют сложную координацию и регуляцию процессов в клетке вследствие заключенной в них генетической информации, обеспечивающей синтез первичной структуры белков-ферментов.
У каждого вида в клетках находится определенное количество хромосом. Они являются носителями генов, определяющих наследственные свойства клеток и организмов вида. Ген – это участок молекулы ДНК хромосомы, на котором синтезируются различные молекулы РНК (трансляторы генетической информации).
В соматических, то есть телесных, клетках обычно содержится двойной, или диплоидный, набор хромосом. Он состоит из пар (2n) практически одинаковых по форме и размеру хромосом. Такие парные, похожие друг на друга хромосомные наборы называют гомологичными (от греч. homos – равный, одинаковый, общий). Они происходят от двух организмов; один набор от материнского, а другой – от отцовского. В таком парном наборе хромосом заключена вся генетическая информация клетки и организма (особи). Гомологичные хромосомы одинаковы по форме, длине, строению, расположению центромеры и несут одни и те же гены, имеющие одинаковую локализацию. Они содержат одинаковый набор генов, хотя и могут различаться их аллелями. Таким образом, гомологичные хромосомы содержат очень близкую, но не идентичную наследственную информацию.
Совокупность признаков хромосом (их число, размеры, форма и детали микроскопического строения) в клетках тела организма того или иного вида называют кариотипом. Форма хромосом, их число, размеры, расположение центромеры, наличие вторичных перетяжек всегда специфичны для каждого вида, по ним можно сопоставлять родство организмов и устанавливать их принадлежность к тому или иному виду.
Постоянство кариотипа, свойственное каждому виду, выработалось в процессе его эволюции и обусловлено закономерностями митоза и мейоза. Однако в процессе существования вида в его кариотипе вследствие мутаций могут произойти изменения хромосом. Некоторые мутации существенно изменяют наследственные качества клетки и организма в целом.
Постоянные характеристики хромосомного набора – количество и морфологические особенности хромосом, определяемые главным образом расположением центромер, наличием вторичных перетяжек, чередованием эухроматиновых и гетерохроматиновых участков и пр., позволяют идентифицировать виды. Поэтому кариотип называют «паспортом» вида .
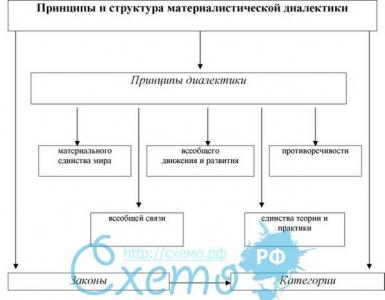
ХРОМОСОМА

СТРОЕНИЕ ХРОМОСОМ
Схема строения хромосомы в поздней профазе - метафазе митоза:
1-хроматида; 2-центромера; 3-короткое плечо; 4-длинное плечо

ЦЕНТРОМЕРА
ЦЕНТРОМЕРА
(от центр + греч. meros - часть) - специализированный участок ДНК, в районе которого в стадии профазы и метафазы деления клетки соединяются две хроматиды, образовавшиеся в результате дупликации хромосомы.

ЗНАЧЕНИЕ ЦЕНТРОМЕРЫ
Центромера играет важную роль при расположении хромосом в виде метафазной пластинки
В процессе расхождения дочерних хромосом к полюсам клетки, так как при помощи центромеры каждая хроматида соединяется с нитями веретена деления.
Каждая центромера разделяет хромосому на два плеча.

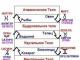
ХРОМАТИДА
ХРОМАТИДА
(от греч. chroma - цвет, краска + eidos - вид) - часть хромосомы от момента ее дупликации до разделения на две дочерние в анафазе, представляет собой нить молекулы ДНК соединенную с белками.
Хроматиды образуются в результате дупликации хромосом в процессе деления клетки.

Хромосомы имеются в ядрах всех клеток.
Каждая хромосома содержит наследственные инструкции - гены.

ГОМОЛОГИЧНЫЕ ХРОМОСОМЫ
От греч.Гомос - одинаковый
Гомологичные хромосомы - парные хромосомы, одинаковые по форме, размерам и набору генов.

ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ
В клетках тела двуполых животных и растений каждая хромосома представлена двумя гомологичными хромосомами, происходящими одна от материнского, а другая от отцовского организма. Такой набор хромосом называют диплоидным
(двойным
)

ГАПЛОИДНЫЙ НАБОР ХРОМОСОМ
Половые клетки, образовавшиеся в результате мейоза, содержат только одну из двух гомологичных хромосом. Этот набор хромосом называют гаплоидным
(одинарным).

ФУНКЦИИ ХРОМОСОМ
Осуществляют координацию и регуляцию процессов в клетке путем синтеза первичной структуры белка, информационной и рибосомальной РНК

ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У РАСТЕНИЙ

ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У ЖИВОТНЫХ
КОМАР – 6 ОКУНЬ – 28 ПЧЕЛА – 32 СВИНЬЯ – 38 МАКАК-РЕЗУС –42 КРОЛИК - 44

24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).

24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).

ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА

3-короткое плечо; 4-длинное плечо
ЦЕНТРОМЕРА
ЦЕНТРОМЕРА
(от центр + греч. meros - часть) - специализированный участок ДНК, в районе которого в стадии профазы и метафазы деления клетки соединяются две хроматиды, образовавшиеся в результате дупликации хромосомы.
ЗНАЧЕНИЕ ЦЕНТРОМЕРЫ
Центромера играет важную роль при расположении хромосом в виде метафазной пластинки
В процессе расхождения дочерних хромосом к полюсам клетки, так как при помощи центромеры каждая хроматида соединяется с нитями веретена деления.
Каждая центромера разделяет хромосому на два плеча.
ХРОМАТИДА
ХРОМАТИДА
(от греч. chroma - цвет, краска + eidos - вид) - часть хромосомы от момента ее дупликации до разделения на две дочерние в анафазе, представляет собой нить молекулы ДНК соединенную с белками.
Хроматиды образуются в результате дупликации хромосом в процессе деления клетки.
Хромосомы имеются в ядрах всех клеток.
Каждая хромосома содержит наследственные инструкции - гены.
ГОМОЛОГИЧНЫЕ ХРОМОСОМЫ
От греч.Гомос - одинаковый
Гомологичные хромосомы - парные хромосомы, одинаковые по форме, размерам и набору генов.
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ
В клетках тела двуполых животных и растений каждая хромосома представлена двумя гомологичными хромосомами, происходящими одна от материнского, а другая от отцовского организма. Такой набор хромосом называют диплоидным
(двойным
)
ГАПЛОИДНЫЙ НАБОР ХРОМОСОМ
Половые клетки, образовавшиеся в результате мейоза, содержат только одну из двух гомологичных хромосом. Этот набор хромосом называют гаплоидным
(одинарным).
ФУНКЦИИ ХРОМОСОМ
Осуществляют координацию и регуляцию процессов в клетке путем синтеза первичной структуры белка, информационной и рибосомальной РНК
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У РАСТЕНИЙ
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У ЖИВОТНЫХ
КОМАР – 6 ОКУНЬ – 28 ПЧЕЛА – 32 СВИНЬЯ – 38 МАКАК-РЕЗУС –42 КРОЛИК - 44
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
ЦЕНТРОМЕРА
ЦЕНТРОМЕРА (от центр + греч. meros - часть) - специализированный участок ДНК, в районе которого в стадии профазы и метафазы деления клетки соединяются две хроматиды, образовавшиеся в результате дупликации хромосомы.
ЗНАЧЕНИЕ ЦЕНТРОМЕРЫ
Центромера играет важную роль при расположении хромосом в виде метафазной пластинки
В процессе расхождения дочерних хромосом к полюсам клетки, так как при помощи центромеры каждая хроматида соединяется с нитями веретена деления.
Каждая центромера разделяет хромосому на два плеча.
ХРОМАТИДА
ХРОМАТИДА (от греч. chroma - цвет, краска + eidos - вид) - часть хромосомы от момента ее дупликации до разделения на две дочерние в анафазе, представляет собой нить молекулы ДНК соединенную с белками.
Хроматиды образуются в результате дупликации хромосом в процессе деления клетки.
Хромосомы имеются в ядрах всех клеток.
Каждая хромосома содержит наследственные инструкции - гены.
ГОМОЛОГИЧНЫЕ ХРОМОСОМЫ
От греч.Гомос - одинаковый
Гомологичные хромосомы - парные хромосомы, одинаковые по форме, размерам и набору генов.
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ
В клетках тела двуполых животных и растений каждая хромосома представлена двумя гомологичными хромосомами, происходящими одна от материнского, а другая от отцовского организма. Такой набор хромосом называют диплоидным (двойным )
ГАПЛОИДНЫЙ НАБОР ХРОМОСОМ
Половые клетки, образовавшиеся в результате мейоза, содержат только одну из двух гомологичных хромосом. Этот набор хромосом называют гаплоидным (одинарным).
ФУНКЦИИ ХРОМОСОМ
Осуществляют координацию и регуляцию процессов в клетке путем синтеза первичной структуры белка, информационной и рибосомальной РНК
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У РАСТЕНИЙ
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ У ЖИВОТНЫХ
КОМАР – 6 ОКУНЬ – 28 ПЧЕЛА – 32 СВИНЬЯ – 38 МАКАК-РЕЗУС –42 КРОЛИК - 44
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
ПЧЕЛА – 32 СВИНЬЯ – 38 МАКАК-РЕЗУС –42 КРОЛИК - 44
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
МАКАК-РЕЗУС –42 КРОЛИК - 44
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС, 2000).
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
Видеоурок 1: Деление клетки. Митоз
Видеоурок 2: Мейоз. Фазы мейоза
Лекция: Клетка - генетическая единица живого. Хромосомы, их строение (форма и размеры) и функции
Клетка - генетическая единица живого
Базовой единицей жизни признана отдельная клетка. Именно на клеточном уровне происходят процессы, отличающие живую материю от неживой. В каждой из клеток хранится и интенсивно используется наследственная информация о химической структуре белков, которые должны синтезироваться в ней и поэтому она называется генетической единицей живого. Даже безъядерные эритроциты на начальных этапах своего существования обладают митохондриями и ядром. Только в зрелом состоянии они не имеют структур для синтеза белка.
На сегодняшний день науке неизвестны клетки, в которых бы не содержалась ДНК или РНК в качестве носителя геномной информации. При отсутствии генетического материала клетка не способна к синтезу белков, а следовательно – жизни.

ДНК имеется не только в ядрах, ее молекулы содержатся в хлоропластах и митохондриях, эти органоиды могут размножаться внутри клетки.
ДНК в клетке находится в виде хромосом – сложных белково-нуклеиновых комплексов. Хромосомы эукариот локализованы в ядре. Каждая из них представляет собой сложную структуру из:
Единственной длинной молекулы ДНК, 2 метра которой упакованы в компактную структуру размером (у человека) до 8 мкм;
Специальных белков-гистонов, чья роль сводится к упаковке хроматина (вещества хромосомы) в знакомую палочковидную форму;
Хромосомы, их строение (форма и размеры) и функции
Такая плотная упаковка генетического материала производится клеткой перед делением. Именно в этот момент можно рассмотреть в микроскоп плотно упакованные сформированные хромосомы. Когда ДНК свернута в компактные хромосомы, называемые гетерохроматином – синтез матричной РНК невозможен. В период набора клеткой массы и ее интерфазного развития хромосомы находятся в менее упакованном состоянии, которое названо интерхроматином и в нем синтезируется мРНК, происходит репликация ДНК.

Основными элементами структуры хромосом являются:
Центромера. Это часть хромосомы с особой последовательностью нуклеотидов. Она соединяет между собой две хроматиды, участвует в конъюгации. Именно к ней прикрепляются белковые нити трубочек веретена деления клетки.
Теломеры . Это концевые участки хромосом, которые не способны к соединению с другими хромосомами, они играют защитную роль. Состоят из повторяющихся участков специализированной ДНК, образующей комплексы с белками.
Точки инициации репликации ДНК.
Хромосомы прокариот очень сильно отличаются от эукариотических, представляя собой расположенные в цитоплазме, ДНК-содержащие структуры. Геометрически они представляют собой кольцевую молекулу.

Хромосомный набор клетки имеет свое название – кариотип. Каждый из видов живых организмов имеет свои, характерные только для него состав, количество и форму хромосом.

Соматические клетки содержат диплоидный (двойной) хромосомный набор, от каждого из родителей получено по половине.
Хромосомы, отвечающие за кодирование одних и тех же функционально белков – называются гомологичными. Плоидность клеток может быть различной – как правило, у животных гаметы гаплоидны. У растений полиплоидия в настоящее время довольно частое явление, использующееся при создании новых сортов в результате гибридизации. Нарушение количества плоидности у теплокровных и человека вызывает серьезные врожденные заболевания, такие как синдром Дауна (наличие трех копий 21-й хромосомы). Чаще всего хромосомные нарушения приводят к нежизнеспособности организма.
У человека полный хромосомный набор состоит из 23 пар. Наибольшее известное число хромосом – 1600, найдено у простейших планктонных организмов – радиолярий. Самый маленький хромосомный набор у австралийских черных муравьев-бульдогов – всего 1.
Жизненный цикл клетки. Фазы митоза и мейоза
Интерфаза , иначе говоря, отрезок времени между двумя делениями, определяется наукой как жизненный цикл клетки.
В течение интерфазы в клетке совершаются жизненные химические процессы, она растет, развивается, накапливает запасные вещества. Подготовка к размножению предусматривает удвоение содержимого – органоидов, вакуолей с питательным содержимым, объема цитоплазмы. Именно благодаря делению, как способу быстрого увеличения количества клеток возможны продолжительная жизнь, размножение, увеличение размеров организма, его выживание при ранениях и регенерация тканей. В клеточном цикле выделяются следующие этапы:
Интерфаза. Время между делениями. Сначала клетка растет, затем увеличивается число органоидов, объем запасного вещества, синтезируются белки. В последней части интерфазы хромосомы готовы к последующему делению – они состоят из пары сестринских хроматид.
Митоз. Так называется один из способов деления ядра, характерный для телесных (соматических) клеток, в его ходе из одной получаются 2 клетки, с идентичным набором генетического материала.
Для гаметогенеза характерен мейоз. Прокариотические клетки сохранили древний способ размножения - прямое деление.
Митоз состоит из 5 основных фаз:
- Анафаза . Усилия микротрубочек отрывают соединения хромосом в области центромеры, и с силой растягивают их к полюсам клетки. При этом хромосомы из-за сопротивления цитоплазмы иногда принимают V-образную форму. В области метафазной пластинки появляется кольцо из белковых волокон.
- Телофаза. Ее началом считается момент, когда хромосомы достигают полюсов деления. Начинается процесс восстановления внутренних мембранных структур клетки – ЭПС, аппарата Гольджи, ядра. Хромосомы распаковываются. Собираются ядрышки, начинается синтез рибосом. Веретено деления распадается.
- Цитокинез . Последняя фаза, в которой появившееся в центральной области клетки белковое кольцо начинает сжиматься, расталкивая цитоплазму к полюсам. Происходит разделение клетки на две и образование на месте белкового кольца клеточной мембраны.
Профаза. Ее началом считается момент, когда хромосомы становятся столь плотно упакованными, что видны в микроскоп. Также, в этом время разрушаются ядрышки, образуется веретено деления. Активизируются микротрубочки, продолжительность их существования уменьшается до 15 секунд, но скорость образования вырастает тоже значительно. Центриоли расходятся к противоположным сторонам клетки, формируя огромное количество постоянно синтезирующихся и распадающихся белковых микротрубочек, которые протягиваются от них к центромерам хромосом. Так формируется веретено деления. Такие мембранные структуры как ЭПС и аппарат Гольджи распадаются на отдельные пузырьки и трубочки, беспорядочно расположенные в цитоплазме. Рибосомы отделяются от мембран ЭПС.
Метафаза . Образуется метафазная пластинка, состоящая из хромосом, уравновешенных в середине клетки усилиями противоположных центриольных микротрубочек, тянущих их каждая в свою сторону. При этом продолжается синтез и распад микротрубочек, своеобразная их «переборка». Эта фаза самая длительная.
Регуляторами процесса митоза являются специфические белковые комплексы. Результатом митотического деления становится пара клеток, обладающих идентичной генетической информацией. В гетеротрофных клетках митоз протекает быстрее, чем в растительных. У гетеротрофов этот процесс может занимать от 30 минут, у растений – 2-3 часа.
Для генерации организмом клеток, имеющих половинное от нормального количество хромосом используется другой механизм деления – мейоз .
Он связан с необходимостью производства половых клеток, у многоклеточных позволяет избежать постоянного увеличения в два раза количества хромосом в последующем поколении и дает возможность получения новых комбинаций аллельных генов. Он отличается количеством фаз, являясь более длительным. Происходящее при этом уменьшение количества хромосом приводит к образованию 4 гаплоидных клеток. Мейоз представляет собой два деления, следующих друг за другом без перерыва.
Определены следующие фазы мейоза:
Профаза I . Гомологичные хромосомы приближаются друг к другу и продольно объединяются. Такое объединение названо конъюгацией. Затем происходит кроссинговер – двойные хромосомы перекрещиваются своими плечами и обмениваются участками.
Метафаза I. Хромосомы разъединяются и занимают места на экваторе клеточного веретена деления, принимая V-образную форму из-за натяжения микротрубочек.
Анафаза I. Гомологичные хромосомы растягиваются микротрубочками к полюсам клетки. Но в отличие от митотического деления, они расходятся целыми, а не отдельными хроматидами.

Результатом первого деления мейоза становится образование двух клеток, имеющих половинное количество целых хромосом. Между делениями мейоза интерфаза практически отсутствует, удвоения хромосом не случается, они и так двуххроматидные.
Сразу следующее за первым повторное мейотическое деление полностью аналогично митозу – в нем хромосомы разделяются на отдельные хроматиды, распределяемые поровну между новыми клетками.
оогонии проходят стадию митотического размножения на эмбриональном этапе развития, так что женский организм уже рождается с неизменным их количеством;
сперматогонии способны к размножению в любой момент репродуктивного периода мужского организма. Генерируется их намного большее количество, чем женских гамет.

Гаметогенез животных организмов происходит в половых железах – гонадах.
Процесс превращения сперматогониев в сперматозоиды происходит в несколько этапов:
Митотическое деление превращает сперматогонии в сперматоциты 1-го порядка.
В результате однократного мейоза они превращаются в сперматоциты 2-го порядка.
Второе мейотическое деление позволяет получить 4 гаплоидные сперматиды.
Наступает период формирования. В клетке происходит уплотнение ядра, уменьшение количества цитоплазмы, формирование жгутика. Также, запасаются белки и увеличивается количество митохондрий.
Формирование яйцеклеток во взрослом женском организме происходит следующим образом:
Из овоцита 1-го порядка, которых в организме находится определенное количество, в результате мейоза с уменьшением количества хромосом вдвое образуются ооциты 2-го порядка.
В результате второго мейотического деления образуются: зрелая яйцеклетка и три мелких редукционных тельца.
Это неравновесное распределение питательных веществ между 4-мя клетками призвано обеспечить большой ресурс питательных веществ для нового живого организма.
Яйцеклетки у папоротников и мхов образуются в архегониях. У более высокоорганизованных растений – в специальных семяпочках, расположенных в завязи.
| | |